B-cells are an integral part of the adaptive immune system. This article delves into B-cell structure, developmental pathways, the dynamic interaction between B-cells and T-cells, and antibody production as part of the immune response against pathogens.
Immunity conferred by B-cells is called humoral immunity, since the B-cells originate and mature in the the bone marrow (the humerus). B-cells are designed to recognise non-self antigens (*1), and produce complementary antibodies in response to the antigen. Thanks to their ability to rearrange genes during maturation, B-cells can, in theory, produce up to 1011 different antibodies in an individual.
B-cell structure
Each B-cell has a B-cell receptor (BCR) on its surface. The BCR is a transmembrane protein that extends into the cytoplasm. During an early stage of development, B-cells undergo rearrangement of the V-DJ genes (1) . V and D/J genes control the sequence of amino acids that make up the tertiary structure of the BCR, and antibodies. Gene rearrangement allows each type of B-cell to produce a unique BCR , and soluble antibodies that have variable regions complementary to only the 3D structure of a one antigen (*2).
B-cells also have co-receptor proteins on their surface. When an antigen is able to cross-link both the BCR and the co-receptor, e.g. CD21, the efficacy of signal protein activation increases multiple-fold (2).
How B-cells work
When they encounter a pathogen or antigen, B-cells process the antigen and present it on the MHC Class II proteins on their surface, ready for presenting to a primed CD4+ T-cells . Crossing-linking of the antigen by the T-cell receptor, and co-receptor CD4 triggers a developmental pathway in CD4+ cells, which mature into CD4+ T-helper cells. (3) Another important molecular interaction during B-cell development is between the CD40 receptor on the B-cells, and the CD40 Ligand on the T-cell – this interaction is important to allow antibody class switching in plasma cells.
Activated CD4+ T-helper cells produce cytokines, including interleukins (IL) and tumour-necrosis factor (TNF alpha) . B-cells have complementary glycoprotein receptors that bind to cytokines, like IL-4 an IL-7, produced by T-helper cells (4). These cytokines play an important role in the differentiation and proliferation of B-cells.
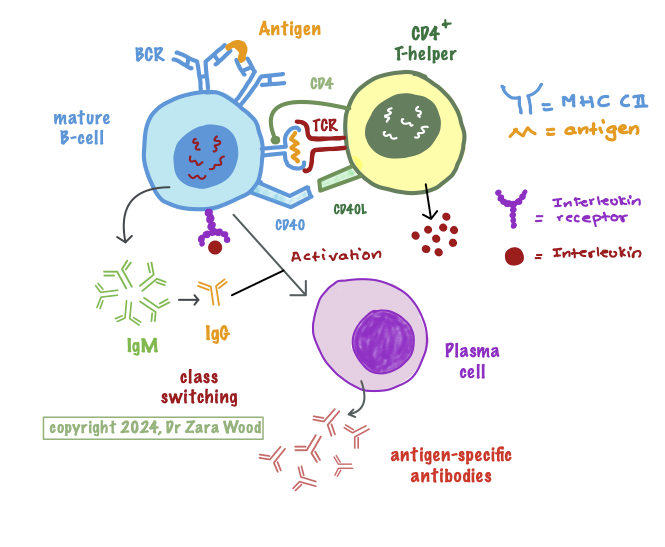
Mature naive B-cells, which have cross-linked with antigen using the BCR, present the antigen to CD4+ T-cells as a peptide-MHC class II complex. CD4 acts as a co-receptor, binding tnonpolymorphic regions on the MHC class II molecule. Class switching is initiated by the interaction of the CD40 ligand on T-cells with the CD40 receptor on B-cells. Naive CD4+ T-cells mature into CD4+ T-helper cells, and further into the TH2 sub-type, which secrete various cytokines like interleukins IL-6 and IL-10. Interleukins attach to receptors on B-cells, which initiates a signal transduction cascade inside the cell, allowing B-cells to clone themselves by mitosis, and divide and differentiate into plasma cells and B-memory cells. Plasma cells produce soluble antibodies, which are antigen/epitope-specific.
What happens when B-cells are activated
Activated B-cells proliferate by mitosis, producing clones of genetically-identical cells. Antigen cross-linking of the BCR, and binding of cytokines to cell-surface receptors on B-cells, activates internal cytoplasmic signal proteins. In turn, signal proteins activate transcription factors, that move to the nucleus and turn on genes responsible for antibody production. This allow antigen-specific B-cells to mature into plasma cells. Each plasma cell produces antibodies of the same antigen specificity.
Each B-cell is capable of producing one of the 5 classes of antibodies – IgA, IgM, IgD, IgG and IgE. A mature B-cell expresses IgM and IgD B-cell receptors, and can produce soluble IgM protein. However, interaction with the CD40 ligand on T-cells allows B-cells to undergo antibody class switching, without the loss of antigen-specificity – the variable region on the antibody remains unchanged, but the constant region changes to a different antibody class.
Switching of antibody class is necessary to allow efficient opsonisation and removal of pathogens by phagocytosis. Class switching allows some plasma cells to switch from IgM to IgG production – IgG is the most abundant antibody in blood plasma. Plasma cells in the mucosa produce IgA whereas IgE is produced in response to an allergen (5).
Primary Immune Response
A primary immune response is the term used to categorise a B-cell response when it has encountered antigen for the first time. Naive mature B-cells undergo antibody class-switching, and start to produce soluble IgG antibodies. These cells are called plasma cells.
Plasma cell numbers rise rapidly in response to an infection, and fall rapidly once infection has been controlled.
Secondary Immune Response
A subset of activated B-cells mature into B-memory cells, which are long-lived. They will respond rapidly to the antigen on subsequent exposure, and generate a secondary immune response that produces large quantities of IgG antibodies. Understanding this response has been important for the development of vaccines.
How antibodies work
Antibodies work by acting as opsonins – their attachment to the surface antigens on the pathogen puts a ‘target’ on the pathogen, allowing a phagocyte to bind to the opsonin (antibody) and ingest the pathogen.
Similarly, antibodies can also cross-link bacteria, in a process called agglutination, that allows for rapid ingestion of pathogens by phagocytes, before they have a chance to multiply and spread around the body. Phagocytes have receptors that bind to the constant region of the antibody, which facilitates ingestion. Once ingested, hydrolytic enzymes released by intra-cellular lysosomes will result in the breakdown of the pathogen.
A similar process occurs to neutralise bacterial toxins. For example, antibodies can neutralise the Cholera toxin or Shiga toxins before they have a chance to bind to complementary receptors on the epithelial cells lining the small intestine. In addition, IgG antibodies can activate the complement cascade, which is an important defence mechanisms of the innate immune system. IgG antibody is the only antibody that can cross the placenta, and provides protection during the first 6 months of life.
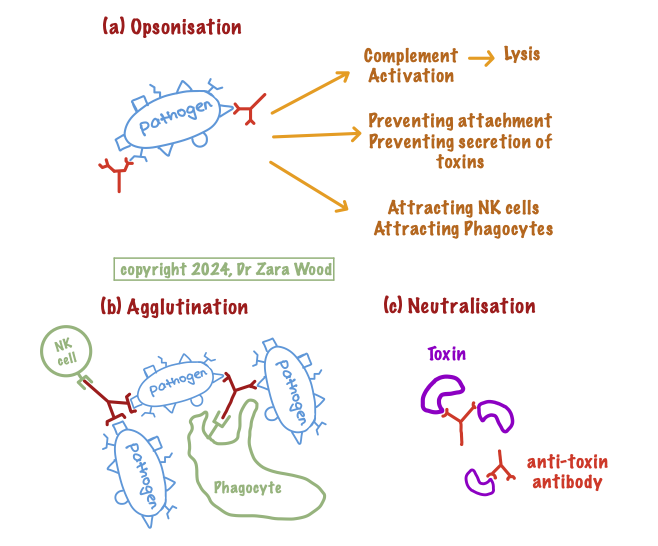
Figure 2: How antibodies work.
(a) Antibodies as Opsonins – antibodies ‘tag’ the antigen on the pathogen through their variable region. This allows phagocytes and NK cells to bind to the antibody via its constant region. The constant region can also activate the complement cascade, a non-specific defence mechanism that results in cell lysis. Opsonins can also block attachment of pathogens to cell-surfaces via their antigen, or prevent secretion of extracellular toxins.
(b) Antibodies in Agglutination – antibodies cross-link antigens on two pathogens, as the two ‘arms’ of the antibody can rotate independently. This causes pathogen cells to ‘clump’ together, making it easier for the phagocyte to ingest and digest large numbers of pathogens.
(c) Antibodies as anti-toxins – anti-toxin antibodies can bind to soluble toxins, and prevent the toxin from binding to, or entering, the cell-surface membrane. They also help with rapid clearance of the toxin from the body.
Other roles of B-cells
Apart from producing antibodies, B-cells also play a key role in immune homeostasis (6). B-cells release cytokines that modulate activity of T-helper and dendritic cells, help with would healing, and the regulation of the inflammatory response. B-cells are therefore critical to the proper functioning of the human immune system.
*1 = during foetal development, B-cells that have self-recognising BCRs undergo apoptosis, which prevents their proliferation. However, if some these B-cells escape the apoptosis signal, or if a BCR develops self-antigen recognition by random mutation, a condition called auto-immune disease can develop.
*2 = it many be able to bind to other epitopes with a low affinity
REFERENCES
- Janeway, CA et al. (2001) – The rearrangement of antigen-receptor gene segments controls lymphocyte development
- Blüml, S et al. (2013) – B-cell targeted therapeutics in clinical development
- Riberdy, JM et al. (1998) – Disruption of the CD4–major histocompatibility complex class II interaction blocks the development of CD4+ T cells in vivo
- Vazquez, MI et al. (2015) – B cells responses and cytokine production are regulated by their immune microenvironment
- Stavnezer, J & Schrader, CE (2014) – Ig heavy chain class switch recombination: mechanism and regulation
- Althwaiqeb, SA & Bordoni,B (2023) – Histology, B Cell Lymphocyte
